
Accueil > 02 - SCIENCES - SCIENCE > Evolution ou révolution des espèces ? > Les arborescences de l’évolution
Les arborescences de l’évolution
lundi 14 janvier 2013, par
Les arborescences de l’évolution

On connait l’image de l’arbre pour l’évolution darwinienne des espèces. L’idée consiste à indiquer l’ancienneté de l’ancêtre commun et à faire figurer cet historique en termes de branches et de sous-branches.

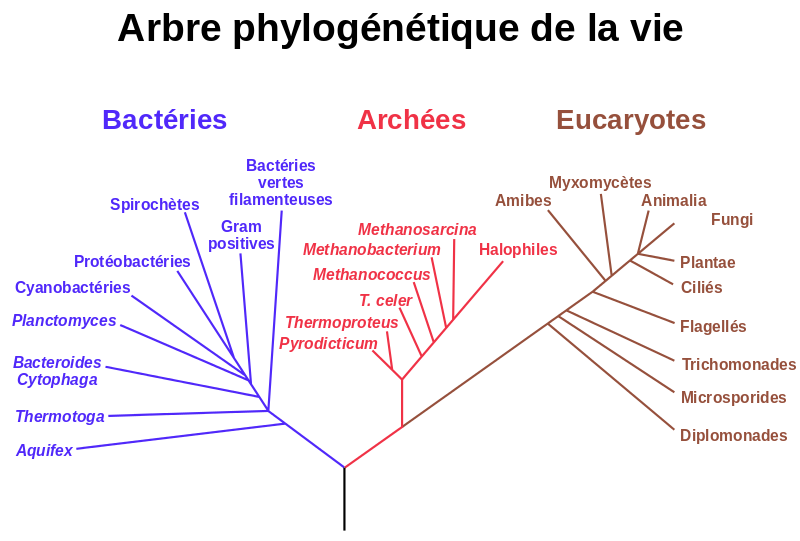
Un arbre phylogénétique est un arbre schématique qui montre donc les relations de parentés entre des groupes d’êtres vivants. Chacun des nœuds de l’arbre représente l’ancêtre commun de ses descendants ; le nom qu’il porte est celui du clade formé des groupes frères qui lui appartiennent, non celui de l’ancêtre qui reste impossible à déterminer. L’arbre peut être enraciné ou pas, selon qu’on est parvenu à identifier l’ancêtre commun à toutes les feuilles. Charles Darwin fut un des premiers scientifiques à proposer une histoire des espèces représentée sous la forme d’un arbre C’est une image maintenant assez ancienne et qui mérite d’être réévaluée.

L’évolution selon Agassiz
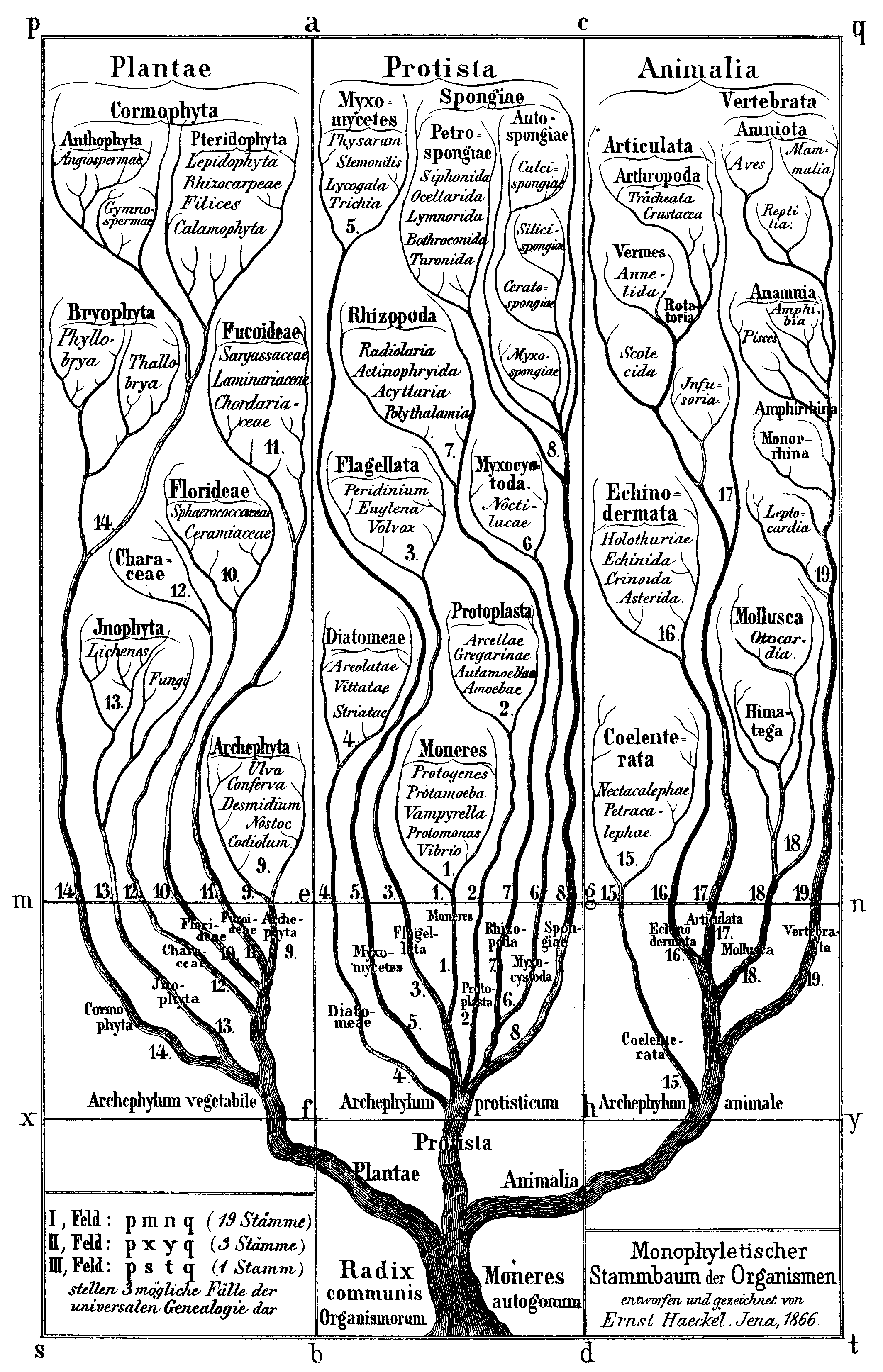
L’image de Haeckel

L’évolution selon Darwin dans First Notebook on Transmutation of Species (1837).
Darwin explique :
« Les affinités de tous les êtres de la même classe ont parfois été représentées sous la forme d’un grand arbre. Je crois que cette comparaison est très juste. Les rameaux verts et bourgeonnants peuvent représenter les espèces existantes ; les branches produites les années précédentes peuvent représenter la longue succession des espèces éteintes. À chaque période de croissance tous les rameaux ont essayé de pousser de tous côtés, de surpasser et de tuer les rameaux et les branches environnantes, de la même manière que les espèces et les groupes d’espèces ont de tout temps vaincu d’autres espèces dans la grande bataille de la vie. Les plus grosses branches se sont divisées en grosses branches, et celles-ci en branches de moins en moins grosses, qui n’étaient autrefois, quand l’arbre était jeune, que des petits rameaux bourgeonnants ; et cette relation entre les anciens bourgeons et les plus récents au moyen de branches ramifiées pourrait bien représenter la classification de toutes les espèces éteintes et vivantes en groupes subordonnés à d’autres groupes. Sur les nombreux rameaux qui florissaient alors que l’arbre n’était qu’un arbuste, seulement deux ou trois, maintenant transformés en grosses branches, survivent encore et portent les autres branches ; ainsi chez les espèces qui vivaient lors des anciennes périodes géologiques, très peu ont laissé des descendants vivants et modifiés. Dès la première croissance de l’arbre, de nombreuses branches ont péri et sont tombées ; et ces branches tombées, de taille variable, peuvent représenter ces ordres, ces familles et ces genres tout entiers, qui n’ont plus de représentants vivants, et que nous ne connaissons qu’à l’état de fossile. De la même façon que nous voyons ici et là une branche menue et isolée, surgissant d’une bifurcation inférieure, et qui, par chance a été favorisée et est encore vivante au sommet de l’arbre, nous voyons occasionnellement un animal tel que l’ornithorynque ou le lépidosirène, qui par de petits détails, connecte grâce à ses affinités deux grandes branches de la vie, et qui apparemment a été sauvé d’une compétition fatale par le fait qu’il vivait dans un habitat protégé. Tout comme les bourgeons produisent par croissance de nouveaux bourgeons, et que ceux-ci, s’ils sont vigoureux, forment des branches et surpassent de tous côtés les branches plus faibles, je crois qu’il en est de même pour le grand « arbre de la vie », qui remplit l’écorce terrestre avec ses branches mortes et brisées, et couvre sa surface avec ses belles ramifications toujours actives. »
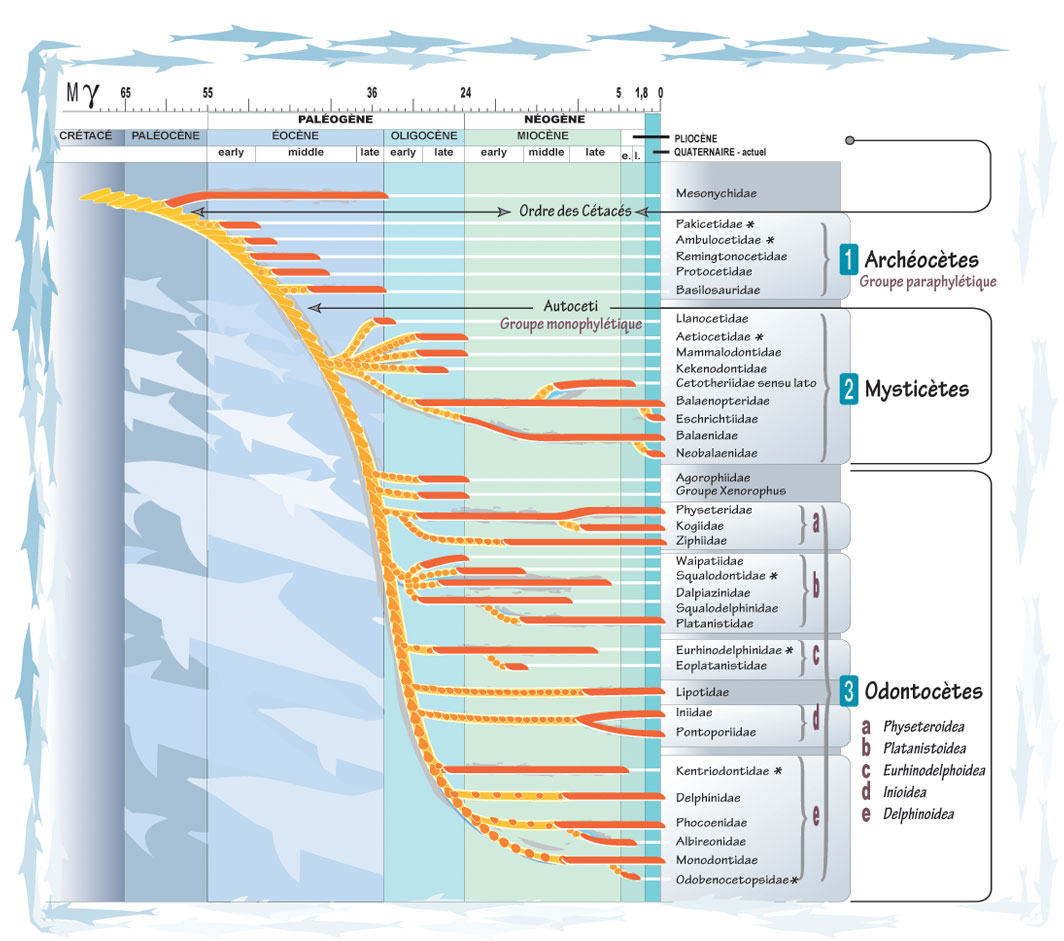
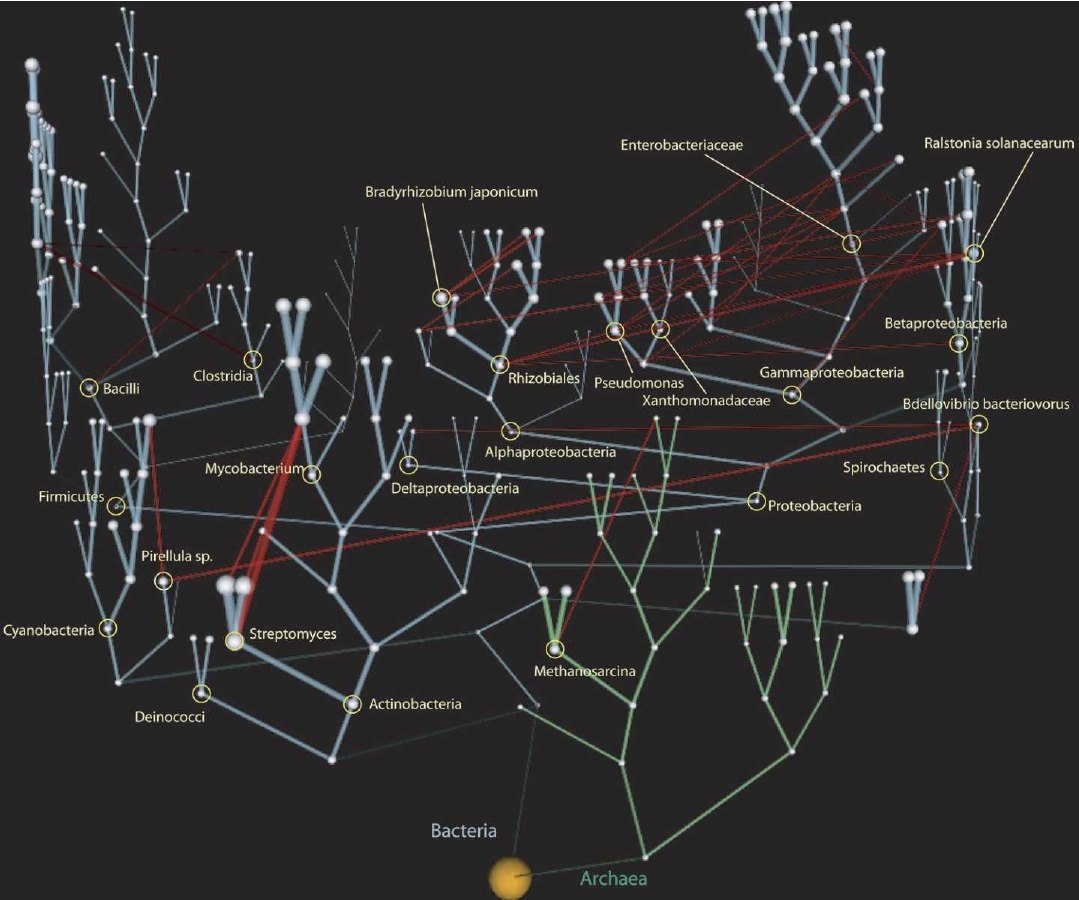

L’évolution en forme d’arbre suppose que les rameaux s’éloignent seulement alors qu’ils s’influencent mutuellement. On assiste à une coévolution et des nouveautés génétiques peuvent passer d’une espèce à une autre.

Mais l’arbre lui-même est une image de continuité puisqu’on passe sans rupture du tronc aux branches et sous-branches. Reste-t-il valable de présenter ainsi l’évolution ? L’arbre n’indique pas le fait qu’une espèce, avant même de donner naissance à de nouvelles espèces, connait une certaine diversification que n’indique pas la stabilité. Il reste aussi le fait que les études indiquent une inquiétante discontinuité dans les fossiles retrouvés.
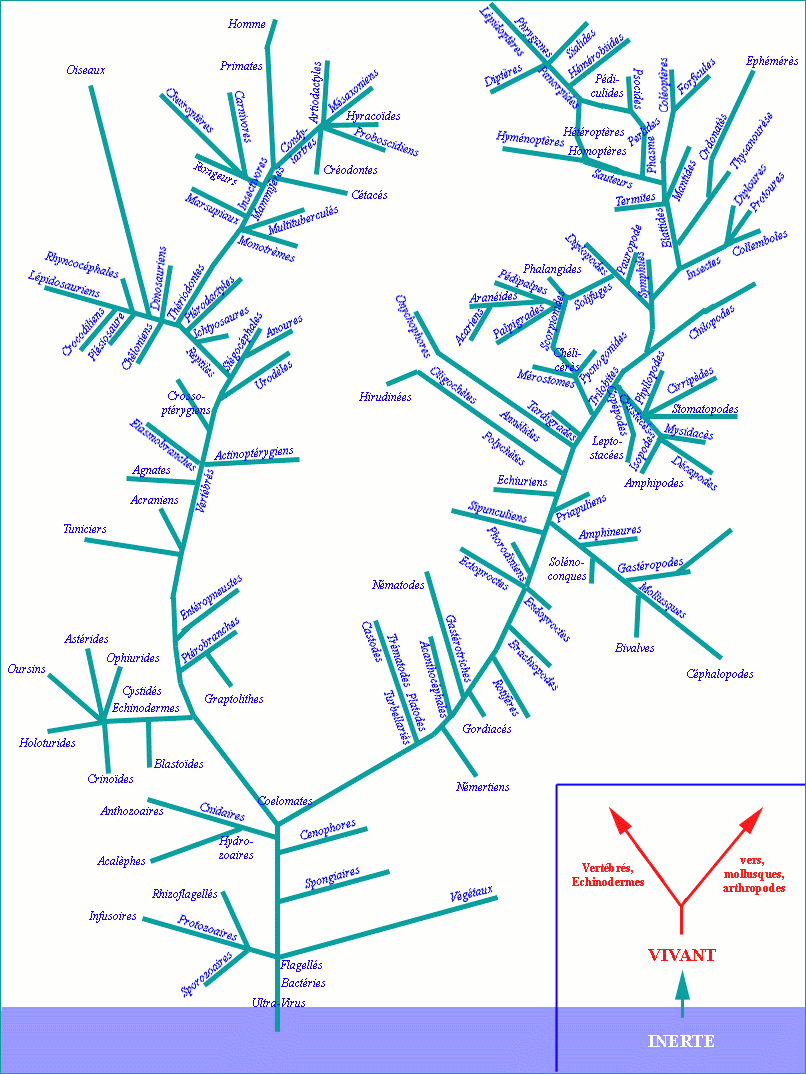
Si on considère l’évolution vers l’homme (expression tout à fait fâcheuse et consciemment) que l’on représente comme un arbre sans être capables en réalité de le reconstituer, on fabrique une arborescence...

... mais on constate que nos observations ne l’indiquent pas mais seulement des segments sans continuité.

On remarque surtout que l’évolution est en buisson qui sont taillés par l’extinction, une image beaucoup plus contradictoire que seulement évolutive.


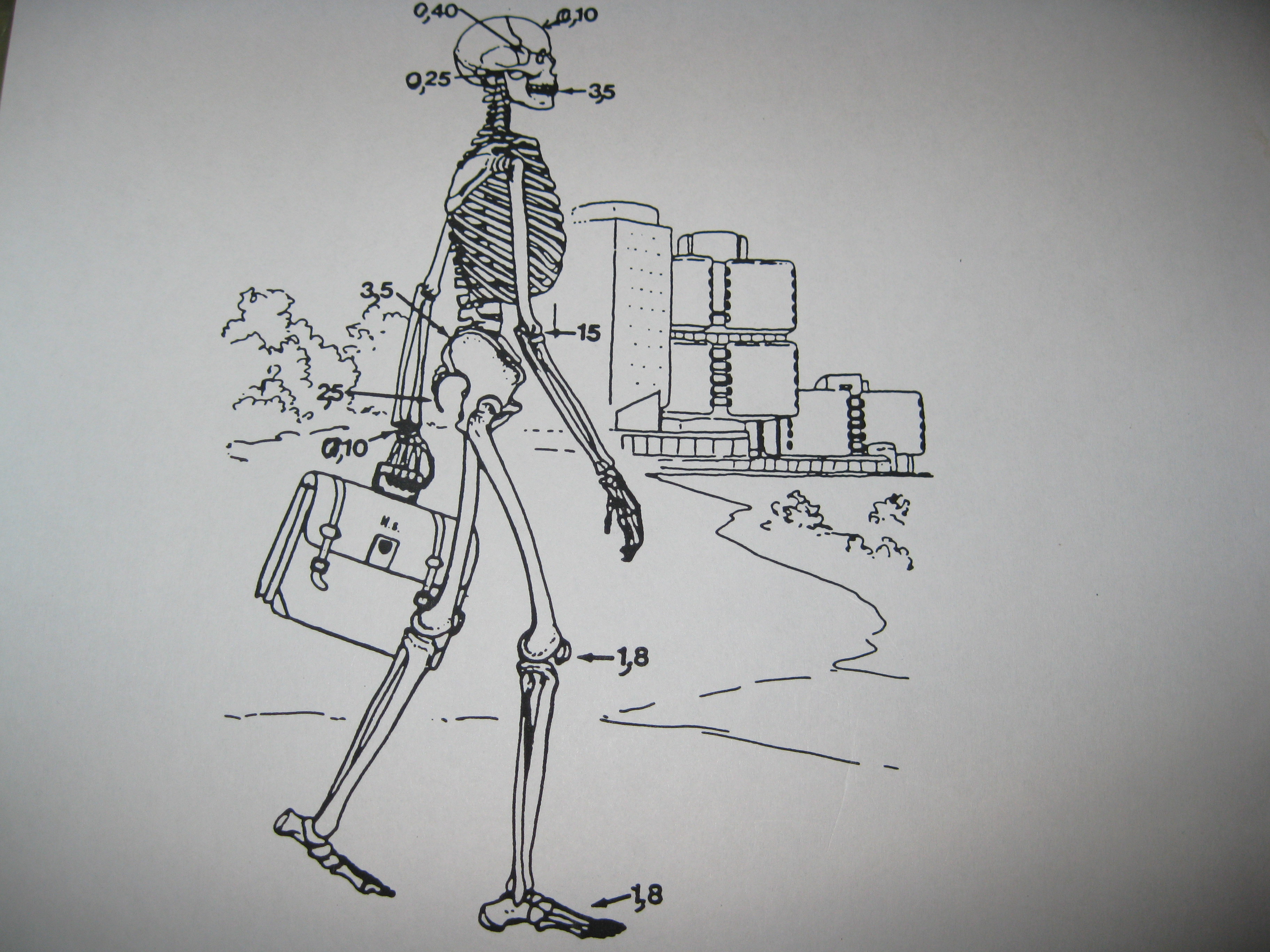
Nous remarquons des patchworks à la manière des anciens châteaux qui contiennent des parties de toutes les époques imbriquées et adaptées les unes aux autres par bricolage. Cela suppose un développement inégal et combiné dans lequel il y a des sauts, des bonds et aussi des sauts dans lesquels un très ancien passé se rappelle au présent…
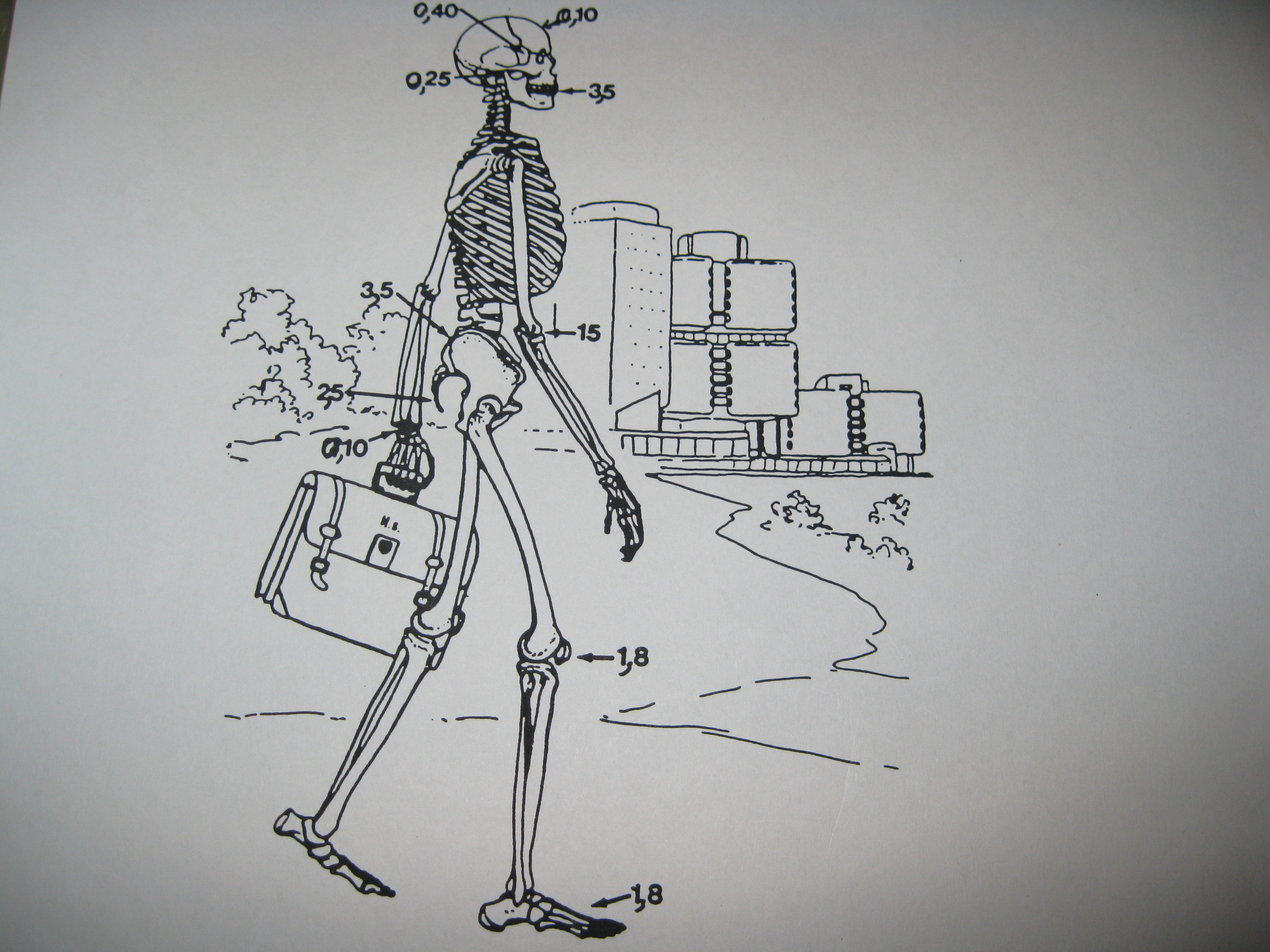
L’évolution est inégale et combinée, ce qui signifie que certaines parties du corps de l’homme sont plus anciennes que d’autres comme un château renaissance qui contient des parties médiévales de différentes époques. L’homme a une formule dentaire acquise par l’évolution il y a 3,5 millions d’années alors que la partie avant de son crâne date de 100.000 ans et la rotule du genou de 1,8 millions d’années.
Donc une espèce n’évolue pas en bloc et les différentes étapes sont des évolutions d’une partie pendant que le reste est inchangé. Cela signifie qu’il n’y a pas une force globale poussant l’ensemble à évoluer… Il y a une diversité naturelle au sein du matériel génétique et un autre mécanisme adverse qui empêche que ces évolutions passent la barrière de vérification des risques de changements. Et parfois cette barrière est affaiblie par une attaque externe (stress) et elle laisse des changements compatibles avec le matériel génétique et même inclus dedans se produire.
L’image de Gould et les équilibres ponctués
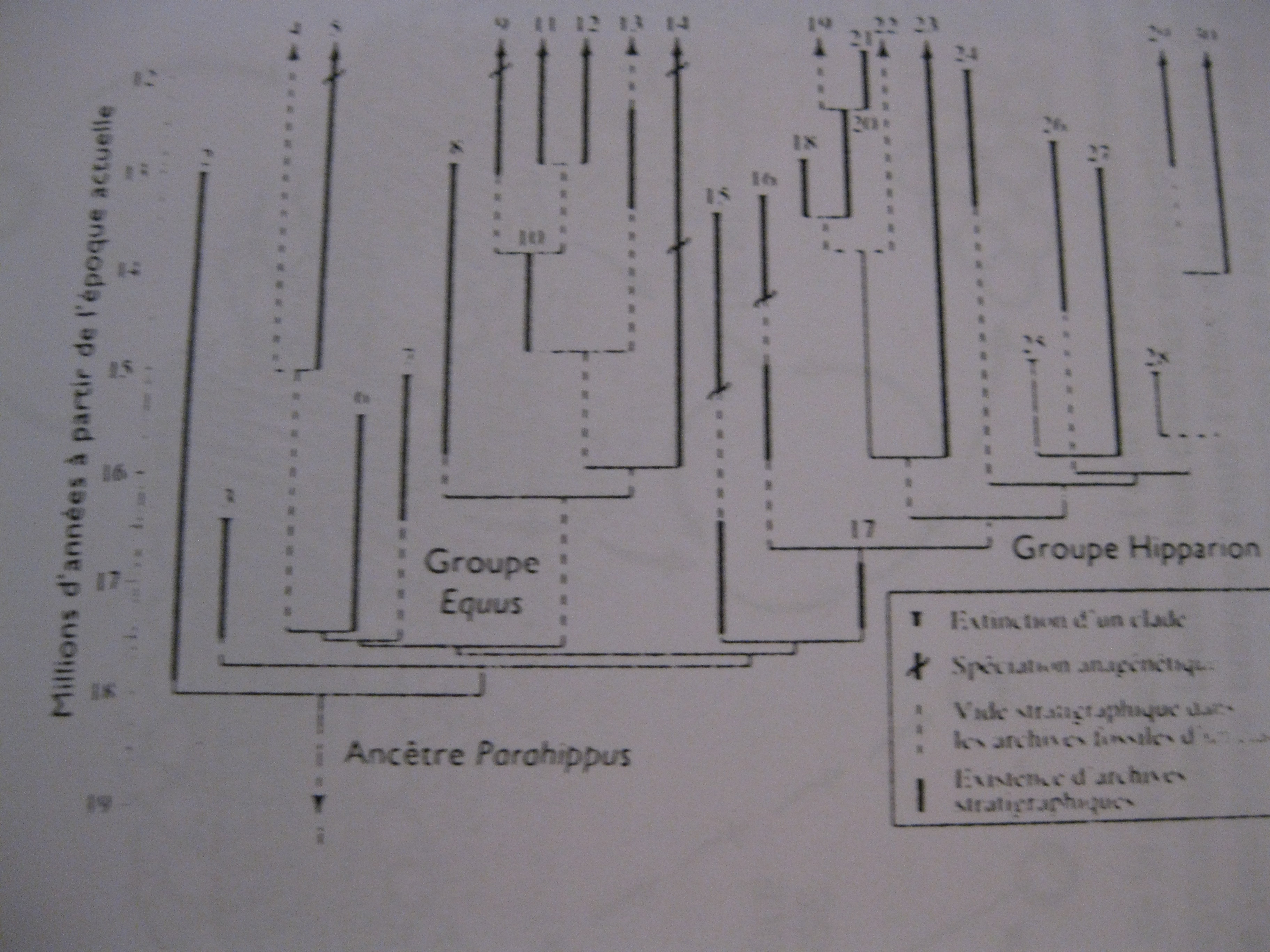
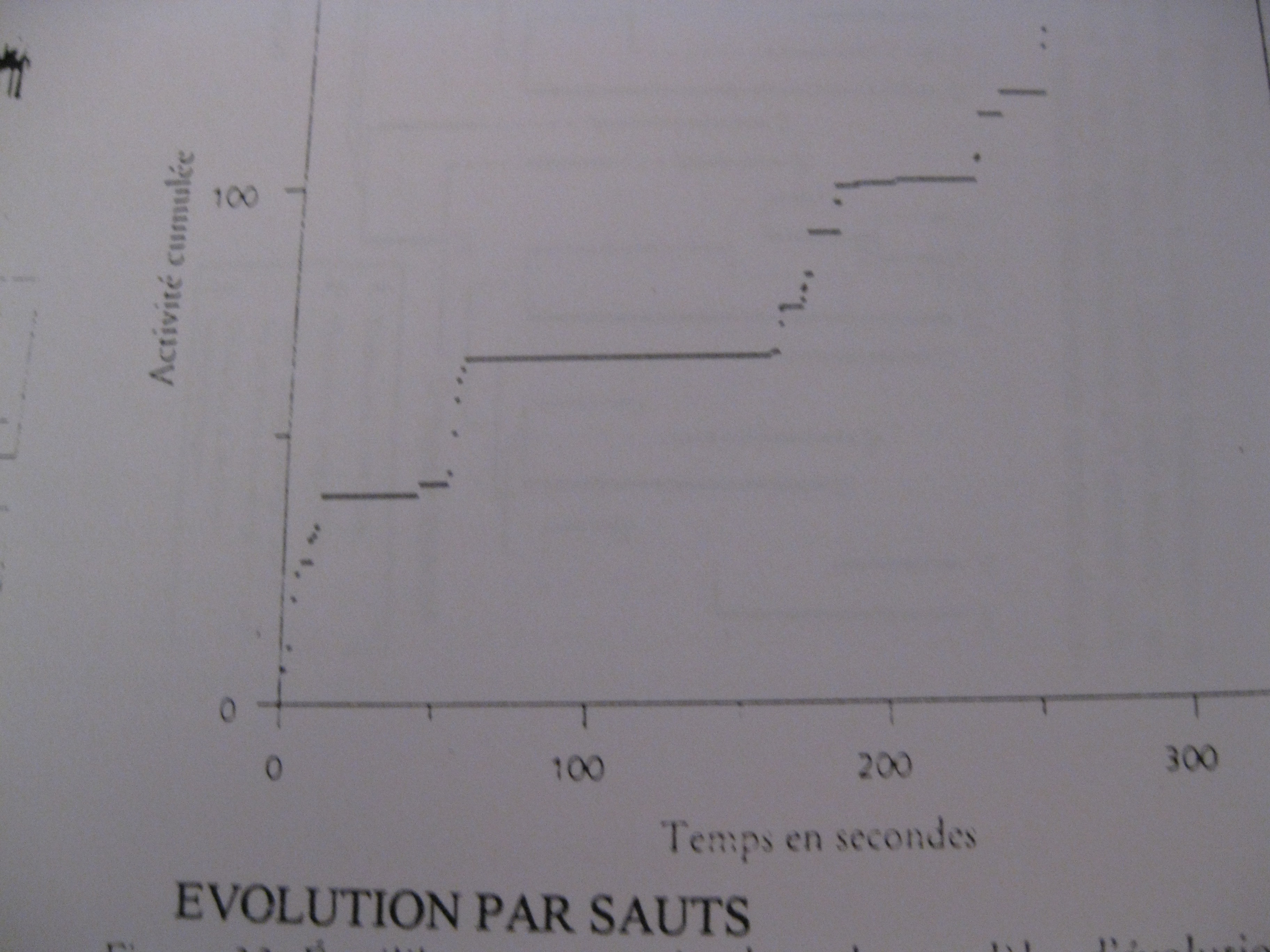
L’évolution n’a rien de continu comme ci-dessous celle de la taille d’un cheval.

D’où l’image des équilibres dits ponctués où l’essentiel du temps il y a équilibre et ponctuellement des sauts.
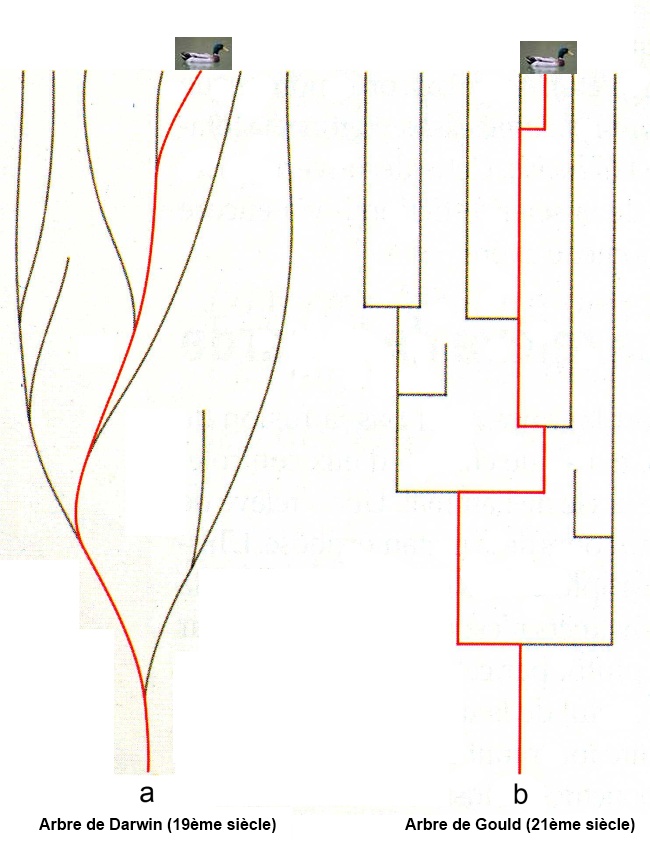
Extrait de Stephen Jay Gould dans "Le pouce du panda" :
"Le caractère épisodique du changement évolutif
Le 23 novembre 1859, le jour précédent la sortie de son livre révolutionnaire, Charles Darwin reçut une lettre extraordinaire de son ami Thomas Henry Huxley. Celui-ci lui offrait son soutien actif dans le combat à venir, allant même jusqu’au sacrifice suprême : « Je suis prêt à mourir sur le bûcher s’il le faut. (…) Je me prépare en aiguisant mes griffes et mon bec. » Mais il ajoutait aussi un avertissement : « Vous vous êtes encombré d’une difficulté inutile en adoptant le « Natura non facit saltum » sans la moindre réserve. » L’expression latine, généralement attribuée à Linné signifie que « la nature ne fait pas de sauts ». Darwin approuvait totalement cette devise ancienne. Disciple de Charles Lyell, apôtre du « gradualisme » en géologie, Darwin décrivait l’évolution comme un processus majestueux et régulier, agissant avec une telle lenteur que personne ne pouvait espérer l’observer pensant la durée d’une vie. Les ancêtres et leurs descendants, selon Darwin, doivent être reliés par « une infinité de liens transitoires » qui forment « une belle succession d’étapes progressives ». Seule une longue période de temps a permis à un processus si lent de réaliser une telle ouvre. Huxley avait le sentiment que Darwin creusait le fossé de sa propre théorie. La sélection naturelle n’avait besoin d’aucun postulat sur la vitesse ; elle pouvait agir tout aussi bien si l’évolution se déroulait sur un rythme rapide. (...)
De nombreux évolutionnistes considèrent qu’une stricte continuité entre micro et macro-évolution constitue un ingrédient essentiel du darwinisme et corollaire nécessaire de la sélection naturelle. (...) Thomas Henry Huxley avait séparé la sélection naturelle du gradualisme et averti Darwin que son adhésion franche et sans fondement sûr au gradualisme pouvait saper son système tout entier. Les fossiles présentent trop de transitions brutales pour témoigner d’un changement progressif et le principe de la sélection naturelle ne l’exige pas, car la sélection peut agir rapidement. Mais ce lien superflu que Darwin a inventé devint le dogme central de la théorie synthétique.
Goldschmidt n’éleva aucune objection contre les thèses classiques de la microévolution. Il consacra la première moitié de son ouvrage principal « Les fondements matériels de l’évolution » au changement progressif et continu au sein des espèces. Cependant, il se démarqua nettement de la théorie synthétique en affirmant que les espèces nouvelles apparaissent soudainement par variation discontinue, ou macro-mutation. Il admit que l’immense majorité des macro-mutations ne pouvaient être considérées que comme désastreuses et il les appela « monstres ». Mais, poursuivit Goldschmidt, une macro-mutation pouvait, par le simple effet de la chance, adapter un organisme à un nouveau mode d’existence. On avait alors affaire, selon sa terminologie, à un « monstre prometteur ». La macro-évolution résulte du succès, peu fréquent, de ces monstres prometteurs, et non de l’accumulation de menus changements au sein des populations. (...) Tous les paléontologistes savent que, parmi les fossiles, on ne compte que peu de formes intermédiaires ; les transitions entre les grands groupes sont particulièrement brutales.
Les gradualistes se sortent habituellement de cette difficulté en invoquant le caractère extrêmement lacunaire des fossiles que nous possédons ; même si une étape sur mille survivait sous forme de fossile, la géologie n’enregistrerait pas le changement continu. (...) Même en l’absence de témoignages directs en faveur de ces transitions sans à-coup peut-on inventer une succession raisonnable de formes intermédiaires, c’est-à-dire des organismes viables, entre les ascendants et les descendants, dans les principales transitions structurelles ? (…)
A quoi sert une moitié de mâchoire et une moitié d’aile ? (...) Si l’on doit accepter de nombreux cas de transition discontinue dans la macroévolution, le darwinisme ne s’effondre-t-il pas en ne survivant que comme une théorie concernant les changements adaptatifs mineurs au sein des espèces ? L’essence même du darwinisme tient en une seule phrase : la sélection naturelle est la principale force créatrice du changement évolutif. Personne ne nie que la sélection naturelle joue un rôle négatif en éliminant les inadaptés. Les théories darwiniennes sous-entendent qu’elle crée en même temps les adaptés. La sélection doit accomplir cette tâche en mettant en place des adaptations en une série d’étapes, tout en préservant à chaque phase le rôle avantageux dans une gamme de variations génétiques dues au hasard.
La sélection doit gouverner le processus de création et non pas se contenter d’écarter les inadaptés après qu’une quelque autre force a soudainement produit une nouvelle espèce complètement achevée dans une perfection primitive. On peut très bien imaginer une théorie non darwinienne du changement discontinu , c’est-à-dire d’une modification génétique profonde et brutale créant par hasard (de temps à autre) et d’un seul coup une nouvelle espèce. Hugo de Vries, le célèbre botaniste hollandais, fut le défenseur de cette théorie. Mais ces notions semblent se heurter à des difficultés insurmontables. (…) Les perturbations apportées aux systèmes génétiques dans leur totalité ne produisent pas de créatures jouissant d’avantages inconnus de leurs descendants – et elles ne sont même pas viables.
Mais toutes les théories du changement discontinu ne sont pas antidarwiniennes, comme l’avait souligné Huxley il y a près de cent vingt ans. Imaginons qu’un changement discontinu dans une forme adulte naisse d’une petite modification génétique. Les problèmes d’incompatibilité avec les autres membres de l’espèce ne se posant pas, cette mutation importante et favorable peut alors se répandre dans la population à la manière darwinienne. Imaginons que ce changement de grande ampleur ne produise pas de suite une forme parfaite, mais serve plutôt d’adaptation clef permettant à son possesseur d’adopter un nouveau modèle d’existence. La poursuite de cette nouvelle vie réussie demande un large ensemble de modifications annexes, tant dans la morphologie que dans le comportement ; ces dernières peuvent survenir en suivant un itinéraire progressif, plus traditionnel, une fois que l’adaptation clef a entraîné une profonde mutation des pressions sélectives.
Les partisans de la synthèse actuelle ont donné à Goldschmidt le rôle de Goldstein en associant son expression imagée – le monstre prometteur – aux notions non darwiniennes de perfection immédiate résultant d’un profond changement génétique. Mais ce n’est pas tout à fait ce que Goldschmidt soutenait En fait, l’un de ses mécanismes entraînant la discontinuité des formes adultes reposait sur la notion de petit changement génétique sous-jacent. Goldschmidt était un spécialiste du développement de l’embryon. Il passa la plus grande partie du début de sa carrière à étudier les variations géographiques de la noctuelle « Lymantria dyspar ». Il découvrit que de grandes différences dans la répartition des couleurs des chenilles provenaient de petits changements dans le rythme du développement : les effets d’un léger retard ou d’un renforcement de la pigmentation au début de la croissance augmentaient à travers l’ontogenèse et entraînaient de profondes différences chez les chenilles ayant atteint leur plein développement. Goldschmidt parvint à identifier les gènes responsables de ces petits changements de rythme et démontra que les grandes différences que l’on observe à la fin du développement proviennent de l’action d’un ou de plusieurs gènes commandant les taux de changement agissant au début de la croissance. Il codifia la notion de « gène de taux de changement » (rate genes) en 1918 et écrivit vingt ans plus tard : « Le gène mutant produit son effet (…) en changeant les taux des processus partiels de développement. Il peut s’agir des taux de croissance ou de différenciation, des taux de production des éléments nécessaires à la différenciation, des taux de réactions entraînant des situations physiques ou chimiques précises à des moments précis du développement, des taux de ces processus responsables de la ségrégation des forces embryonnaires à des moments donnés. » (…)
Selon ma propre opinion, très partiale, le problème de la réconciliation entre l’évidente discontinuité de la macro-évolution et le darwinisme est en grande partie résolu si l’on observe que les changements de faible ampleur survenant tôt dans le développement de l’embryon s’accumulent pendant la croissance pour produire de profondes différences chez l’adulte. En prolongeant dans la petite enfance le rythme élevé de la croissance prénatale du cerveau du singe, on voit sa taille se rapprocher de celle du cerveau humain. (...) En réalité, si l’on n’invoque pas le changement discontinu par de petites modifications dans les taux de développement, je ne vois pas comment peuvent s’accomplir la plupart des principales transitions de l’évolution. » Peu de systèmes présentent une résistance plus grande au changement que les adultes complexes, fortement différenciés, des animaux « supérieurs ». Comment pourrait-on convertir un rhinocéros adulte ou un moustique en quelque chose de foncièrement différent ? Cependant les transitions entre les groupes principaux se sont bien produites au cours de l’histoire de la vie.
D’Arcy Wentworth Thompson (…) écrit dans « Croissance et forme » : « (...) Nous ne pouvons pas transformer un invertébré en vertébré, ni un cœlentéré en vert, par n’importe déformation simple et légitime (…) La nature passe d’un type à un autre. (…) Chercher des marchepieds pour franchir les écarts séparant ces types, c’est chercher en vain à jamais. » La solution de D’Arcy Wentworth Thompson était la même que celle de Goldschmidt : la transition peut se produire dans les embryons qui sont plus simples et plus semblables entre eux que les adultes fortement divergents qu’ils forment. Personne ne songerait à transformer une étoile de mer en souris, mais les embryons de certains échinodermes et de certains protovertébrés sont presque identiques. »



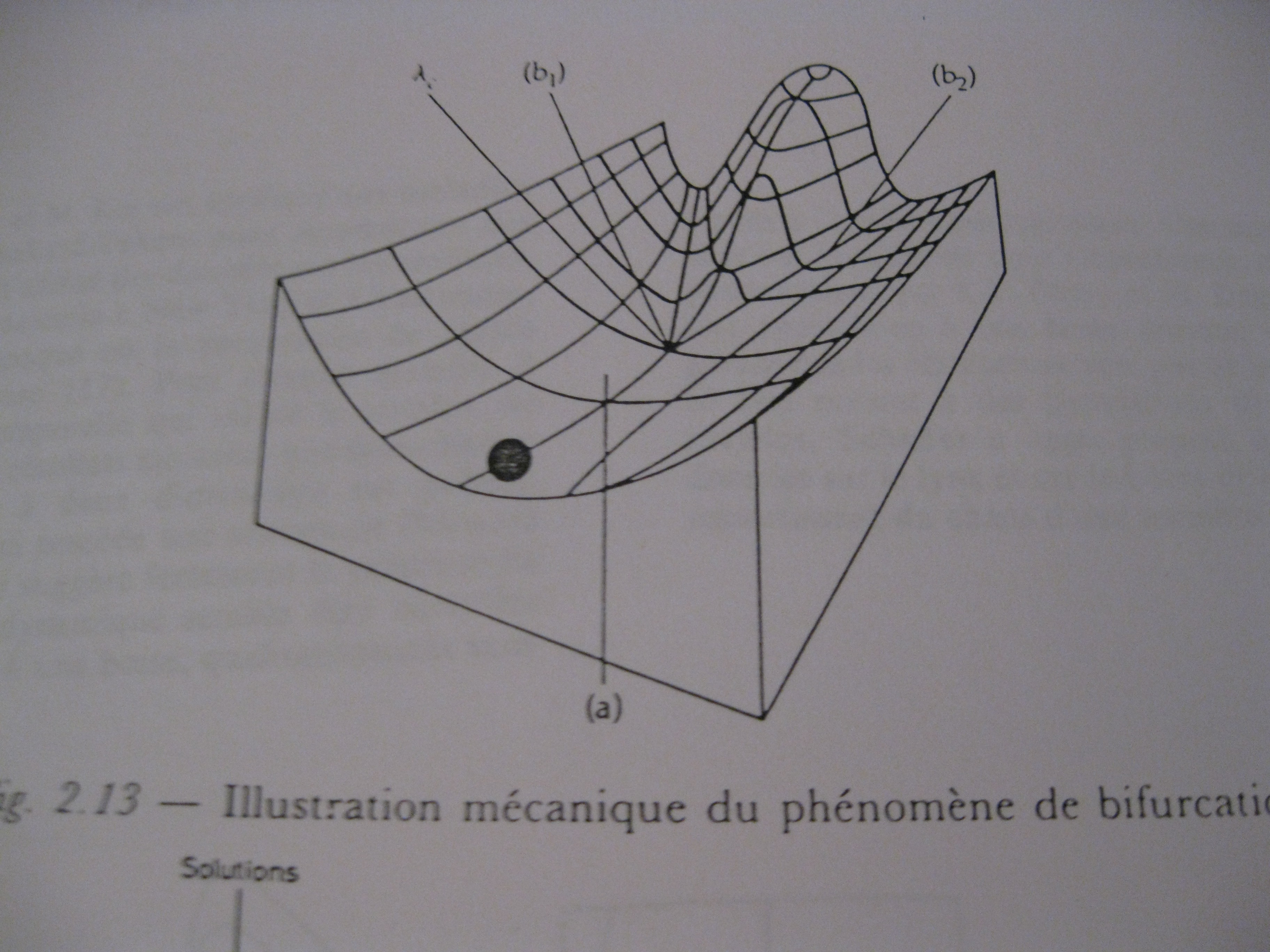
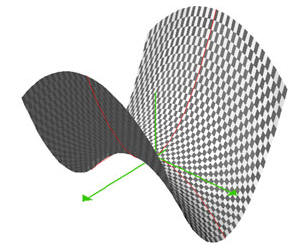
Quelle topologie pour l’espèce sur un palier ? La selle de cheval !

La selle de cheval, ou col, est une figure topologique qui a la particularité d’être une zone dans laquelle la dynamique peut durer très longtemps, y avoir de multiples trajectoire ressemblant à la stabilité et, brutalement, passer à une trajectoire qui fait descendre du col. Et il y a plusieurs manières très divergentes de descendre d’un col... Donc plusieurs évolutions possibles !
Deux mécanismes contradictoires et rétroactifs qui, en explorant le champ des possibles, en passant par plusieurs paliers transitoires, vont finir par trouver un niveau d’équilibre, mais on ne peut pas prévoir à quel moment cet équilibre sera brutalement rompu. Par exemple, la dynamique peut rester très longtemps dans une zone appelée selle de cheval (ou col) en étant sans cesse en transformation mais en conservant des paramètres ne quittant pas cette zone. Elle peut durer longtemps et sembler stable. Puis, tout à coup, du fait d’un petit changement, les boucles de rétroactions sont susceptibles de se désordonner et même de sauter rapidement à un autre niveau d’équilibre. Le col est durable mais il n’est pas stable. Un mécanisme de régulation de ce type peut être construit spontanément par des rétroactions opposées qui explorent tous les modes durables.

L’évolution des espèces vivantes que l’on a cherché à tout prix à imaginer sous forme linéaire du type de cause à effet, comme le singe est cause de l’homme. Mais on s’est, du coup, aperçus que ce paradigme ne fonctionnait pas. Les chaînons manquant se sont multipliés. L’étude de la génétique a souligné des contradictions irréconciliables avec l’image de la causalité linéaire. Il a fallu reconnaître le mode arborescent de l’évolution. Il n’y a pas lien direct de cause à effet entre les espèces que l’on a trouvé, pas plus pour les hominoïdes que pour les autres espèces vivantes. La raison en est que l’on cherche, par erreur, à relier par des segments évolutifs des espèces entièrement distinctes et on efface ainsi la phase transitoire qui a permis cette évolution, phase pendant laquelle les espèces diverses qui ont la même origine ne sont pas encore complètement séparées, restent interfécondes, échangent encore des gènes, s’influencent mutuellement. Cela signifie qu’à un moment, une potentialité de diversité (qui existait déjà potentiellement ce qui signifie que nonA existait au sein de A) se met à s’exprimer dans des sens divers. La vie se met brutalement à explorer la diversité. Mais elle commence par le faire à un stade où la divergence entre les espèces n’est pas encore réalisée. C’est ce qu’en topologie de la dynamique du développement on appelle un col. Les trajectoires des états explorent l’ensemble du col, peuvent y rester assez longtemps, s’entrecroiser, bien avant qu’une trajectoire les fasse quitter le col pour descendre une pente (suivant ainsi un attracteur pour se spécialiser, se distinguer définitivement des autres espèces).

Cette image des arborescences avec divergence et rupture est très différente de celle, linéaire et continue, sans rupture, de « cause à effet ». C’est un faisceau de causes et de circonstances (y compris de hasards) qui entraine un stade de transformation qui culmine dans un changement brutal (ou plusieurs). Le changement est une négation ou une destruction de l’état ancien et pas seulement une conséquence ou une implication (comme dans « cause à effet »).
Chaque évolution définitive est une rupture avec les multiples potentialités qui existaient encore au col. Réaliser une potentialité, c’est donc rompre avec beaucoup d’autres, y renoncer définitivement. C’est cette rupture qui rend l’évolution irréversible et historique alors que la « cause à effet » n’est pas véritablement historique, n’étant fondée sur aucune rupture. C’est cette rupture avec l’arborescence des possibles qui produit de la nouveauté. Le nouvel état se fonde sur un capital différent du tronc de départ et produit des structures nouvelles. Le processus de création n’est donc pas positif, « pour construire », mais négatif, « parce que l’on a détruit et qu’il faut bâtir du neuf ». Les structures nouvelles se succèdent mais pas le long d’une ligne du progrès, au travers des destructions et refondations. La destruction est indispensable à l’évolution. La cause en est profonde : une évolution irréversible suppose la destruction des potentialités, une bifurcation qui est rupture avec le tronc principal sans retour possible.

Messages
1. Les arborescences de l’évolution, 14 janvier 2013, 16:57, par Abdel
Pouvez-vous résumer comment a évolué la conception de l’arbre de la vie ?